Orobanchol
General
Type : Plant hormone || Strigolactone || Strigolactone receptors ligand || Natural
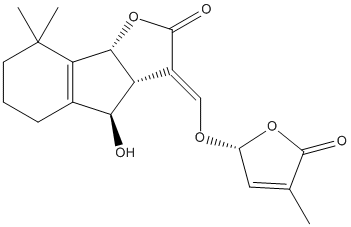
Chemical_Nomenclature : (3E,3aS,4S,8bS)-4-hydroxy-8,8-dimethyl-3-[[(2R)-4-methyl-5-oxo-2H-furan-2-yl]oxymethylidene]-3a,4,5,6,7,8b-hexahydroindeno[1,2-b]furan-2-one
Canonical SMILES : CC1=CC(OC1=O)OC=C2C3C(C4=C(C3OC2=O)C(CCC4)(C)C)O
InChI : InChI=1S\/C19H22O6\/c1-9-7-12(24-17(9)21)23-8-11-13-15(20)10-5-4-6-19(2,3)14(10)16(13)25-18(11)22\/h7-8,12-13,15-16,20H,4-6H2,1-3H3\/b11-8+\/t12-,13+,15-,16+\/m1\/s1
InChIKey : CDBBMEYPRMUMTR-RZXXLYMMSA-N
Other name(s) : (+)-orobanchol, CHEMBL2270919, SCHEMBL13857704
Target
Families : RsbQ-like, Plant_carboxylesterase
References (13)
| Title : Structure Elucidation and Biosynthesis of Orobanchol - Wakabayashi_2022_Front.Plant.Sci_13_835160 |
| Author(s) : Wakabayashi T , Ueno K , Sugimoto Y |
| Ref : Front Plant Sci , 13 :835160 , 2022 |
| Abstract : Wakabayashi_2022_Front.Plant.Sci_13_835160 |
| ESTHER : Wakabayashi_2022_Front.Plant.Sci_13_835160 |
| PubMedSearch : Wakabayashi_2022_Front.Plant.Sci_13_835160 |
| PubMedID: 35222492 |
| Title : Characterization of a Chickpea Mutant Resistant to Phelipanche aegyptiaca Pers. and Orobanche crenata Forsk - Galili_2021_Plants.(Basel)_10_ |
| Author(s) : Galili S , Hershenhorn J , Smirnov E , Yoneyama K , Xie X , Amir-Segev O , Bellalou A , Dor E |
| Ref : Plants (Basel) , 10 : , 2021 |
| Abstract : Galili_2021_Plants.(Basel)_10_ |
| ESTHER : Galili_2021_Plants.(Basel)_10_ |
| PubMedSearch : Galili_2021_Plants.(Basel)_10_ |
| PubMedID: 34961023 |
| Title : Catabolism of strigolactones by a carboxylesterase - Xu_2021_Nat.Plants_7_1495 |
| Author(s) : Xu E , Chai L , Zhang S , Yu R , Zhang X , Xu C , Hu Y |
| Ref : Nat Plants , 7 :1495 , 2021 |
| Abstract : Xu_2021_Nat.Plants_7_1495 |
| ESTHER : Xu_2021_Nat.Plants_7_1495 |
| PubMedSearch : Xu_2021_Nat.Plants_7_1495 |
| PubMedID: 34764442 |
| Gene_locus related to this paper: arath-CXE15 |
| Title : A new UHPLC-MS\/MS method for the direct determination of strigolactones in root exudates and extracts - Rial_2019_Phytochem.Anal_30_110 |
| Author(s) : Rial C , Varela RM , Molinillo JMG , Lopez-Raez JA , Macias FA |
| Ref : Phytochem Anal , 30 :110 , 2019 |
| Abstract : Rial_2019_Phytochem.Anal_30_110 |
| ESTHER : Rial_2019_Phytochem.Anal_30_110 |
| PubMedSearch : Rial_2019_Phytochem.Anal_30_110 |
| PubMedID: 30280444 |
| Title : Direct conversion of carlactonoic acid to orobanchol by cytochrome P450 CYP722C in strigolactone biosynthesis - Wakabayashi_2019_Sci.Adv_5_eaax9067 |
| Author(s) : Wakabayashi T , Hamana M , Mori A , Akiyama R , Ueno K , Osakabe K , Osakabe Y , Suzuki H , Takikawa H , Mizutani M , Sugimoto Y |
| Ref : Sci Adv , 5 :eaax9067 , 2019 |
| Abstract : Wakabayashi_2019_Sci.Adv_5_eaax9067 |
| ESTHER : Wakabayashi_2019_Sci.Adv_5_eaax9067 |
| PubMedSearch : Wakabayashi_2019_Sci.Adv_5_eaax9067 |
| PubMedID: 32064317 |
| Title : Low strigolactone root exudation: a novel mechanism of broomrape (Orobanche and Phelipanche spp.) resistance available for faba bean breeding - Fernandez-Aparicio_2014_J.Agric.Food.Chem_62_7063 |
| Author(s) : Fernandez-Aparicio M , Kisugi T , Xie X , Rubiales D , Yoneyama K |
| Ref : Journal of Agricultural and Food Chemistry , 62 :7063 , 2014 |
| Abstract : Fernandez-Aparicio_2014_J.Agric.Food.Chem_62_7063 |
| ESTHER : Fernandez-Aparicio_2014_J.Agric.Food.Chem_62_7063 |
| PubMedSearch : Fernandez-Aparicio_2014_J.Agric.Food.Chem_62_7063 |
| PubMedID: 24974726 |
| Title : Confirming stereochemical structures of strigolactones produced by rice and tobacco - Xie_2013_Mol.Plant_6_153 |
| Author(s) : Xie X , Yoneyama K , Kisugi T , Uchida K , Ito S , Akiyama K , Hayashi H , Yokota T , Nomura T |
| Ref : Mol Plant , 6 :153 , 2013 |
| Abstract : Xie_2013_Mol.Plant_6_153 |
| ESTHER : Xie_2013_Mol.Plant_6_153 |
| PubMedSearch : Xie_2013_Mol.Plant_6_153 |
| PubMedID: 23204500 |
| Title : A petunia ABC protein controls strigolactone-dependent symbiotic signalling and branching - Kretzschmar_2012_Nature_483_341 |
| Author(s) : Kretzschmar T , Kohlen W , Sasse J , Borghi L , Schlegel M , Bachelier JB , Reinhardt D , Bours R , Bouwmeester HJ , Martinoia E |
| Ref : Nature , 483 :341 , 2012 |
| Abstract : Kretzschmar_2012_Nature_483_341 |
| ESTHER : Kretzschmar_2012_Nature_483_341 |
| PubMedSearch : Kretzschmar_2012_Nature_483_341 |
| PubMedID: 22398443 |
| Title : Ent-2'-epi-Orobanchol and its acetate, as germination stimulants for Striga gesnerioides seeds isolated from cowpea and red clover - Ueno_2011_J.Agric.Food.Chem_59_10485 |
| Author(s) : Ueno K , Nomura S , Muranaka S , Mizutani M , Takikawa H , Sugimoto Y |
| Ref : Journal of Agricultural and Food Chemistry , 59 :10485 , 2011 |
| Abstract : Ueno_2011_J.Agric.Food.Chem_59_10485 |
| ESTHER : Ueno_2011_J.Agric.Food.Chem_59_10485 |
| PubMedSearch : Ueno_2011_J.Agric.Food.Chem_59_10485 |
| PubMedID: 21899364 |
| Title : Strigolactones are transported through the xylem and play a key role in shoot architectural response to phosphate deficiency in nonarbuscular mycorrhizal host Arabidopsis - Kohlen_2011_Plant.Physiol_155_974 |
| Author(s) : Kohlen W , Charnikhova T , Liu Q , Bours R , Domagalska MA , Beguerie S , Verstappen F , Leyser O , Bouwmeester H , Ruyter-Spira C |
| Ref : Plant Physiol , 155 :974 , 2011 |
| Abstract : Kohlen_2011_Plant.Physiol_155_974 |
| ESTHER : Kohlen_2011_Plant.Physiol_155_974 |
| PubMedSearch : Kohlen_2011_Plant.Physiol_155_974 |
| PubMedID: 21119045 |
| Title : Aromatic A-ring analogues of orobanchol, new germination stimulants for seeds of parasitic weeds - Malik_2011_Org.Biomol.Chem_9_2286 |
| Author(s) : Malik H , Kohlen W , Jamil M , Rutjes FP , Zwanenburg B |
| Ref : Org Biomol Chem , 9 :2286 , 2011 |
| Abstract : Malik_2011_Org.Biomol.Chem_9_2286 |
| ESTHER : Malik_2011_Org.Biomol.Chem_9_2286 |
| PubMedSearch : Malik_2011_Org.Biomol.Chem_9_2286 |
| PubMedID: 21321762 |
| Title : A small-molecule screen identifies new functions for the plant hormone strigolactone - Tsuchiya_2010_Nat.Chem.Biol_6_741 |
| Author(s) : Tsuchiya Y , Vidaurre D , Toh S , Hanada A , Nambara E , Kamiya Y , Yamaguchi S , McCourt P |
| Ref : Nat Chemical Biology , 6 :741 , 2010 |
| Abstract : Tsuchiya_2010_Nat.Chem.Biol_6_741 |
| ESTHER : Tsuchiya_2010_Nat.Chem.Biol_6_741 |
| PubMedSearch : Tsuchiya_2010_Nat.Chem.Biol_6_741 |
| PubMedID: 20818397 |
| Title : Strigolactones, host recognition signals for root parasitic plants and arbuscular mycorrhizal fungi, from Fabaceae plants - Yoneyama_2008_New.Phytol_179_484 |
| Author(s) : Yoneyama K , Xie X , Sekimoto H , Takeuchi Y , Ogasawara S , Akiyama K , Hayashi H |
| Ref : New Phytol , 179 :484 , 2008 |
| Abstract : Yoneyama_2008_New.Phytol_179_484 |
| ESTHER : Yoneyama_2008_New.Phytol_179_484 |
| PubMedSearch : Yoneyama_2008_New.Phytol_179_484 |
| PubMedID: 19086293 |