Arachidonoyl-1-thioglycerol
General
Type : Acyl-Glycerol || Monoacylglycerol || Fatty acid ester || Water-insoluble medium and long chain ester
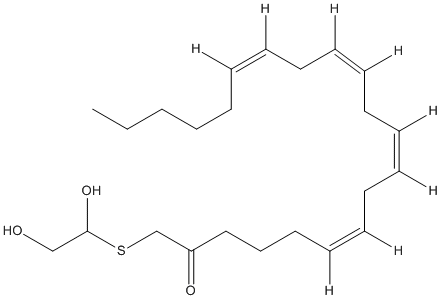
Chemical_Nomenclature : S-(2,3-dihydroxypropyl) (5Z,8Z,11Z,14Z)-icosa-5,8,11,14-tetraenethioate
Canonical SMILES : CCCCC\/C=CC\/C=CC\/C=CC\/C=CCCCC(=O)SCC(CO)O || CCCCCC=CCC=CCC=CCC=CCCCC(=O)SCC(CO)O
InChI : InChI=1S\/C23H38O3S\/c1-2-3-4-5-6-7-8-9-10-11-12-13-14-15-16-17-18-19-23(26)27-21-22(25)20-24\/h6-7,9-10,12-13,15-16,22,24-25H,2-5,8,11,14,17-21H2,1H3\/b7-6-,10-9-,13-12-,16-15- || InChI=1S\/C23H38O3S\/c1-2-3-4-5-6-7-8-9-10-11-12-13-14-15-16-17-18-19-23(26)27-21-22(25)20-24\/h6-7,9-10,12-13,15-16,22,24-25H,2-5,8,11,14,17-21H2,1H3
InChIKey : UUOPRYPOAXYNLX-DOFZRALJSA-N || UUOPRYPOAXYNLX-UHFFFAOYSA-N
Other name(s) : SCHEMBL17046826, HMS3650M03, 1625AH, Arachidonyl-1-thio-glycerol, CTK8F7813, S-(2,3-dihydroxypropyl) icosa-5,8,11,14-tetraenethioate
References (3)
| Title : The Antimalarial Natural Product Salinipostin A Identifies Essential alpha\/beta Serine Hydrolases Involved in Lipid Metabolism in P. falciparum Parasites - Yoo_2020_Cell.Chem.Biol_27_143 |
| Author(s) : Yoo E , Schulze CJ , Stokes BH , Onguka O , Yeo T , Mok S , Gnadig NF , Zhou Y , Kurita K , Foe IT , Terrell SM , Boucher MJ , Cieplak P , Kumpornsin K , Lee MCS , Linington RG , Long JZ , Uhlemann AC , Weerapana E , Fidock DA , Bogyo M |
| Ref : Cell Chemical Biology , 27 :143 , 2020 |
| Abstract : Yoo_2020_Cell.Chem.Biol_27_143 |
| ESTHER : Yoo_2020_Cell.Chem.Biol_27_143 |
| PubMedSearch : Yoo_2020_Cell.Chem.Biol_27_143 |
| PubMedID: 31978322 |
| Gene_locus related to this paper: plaf7-q8ii19 , plafa-a0a143zya4 , plaf7-q8iik5 , plafa-MAL8P1.38 , plafa-PF07.0040 , plafa-PF10.0020 , plafa-PF10.0379 , plafa-PF13.0153 |
| Title : Simplified assays of lipolysis enzymes for drug discovery and specificity assessment of known inhibitors - Iglesias_2016_J.Lipid.Res_57_131 |
| Author(s) : Iglesias J , Lamontagne J , Erb H , Gezzar S , Zhao S , Joly E , Truong VL , Skorey K , Crane S , Madiraju SR , Prentki M |
| Ref : J Lipid Res , 57 :131 , 2016 |
| Abstract : Iglesias_2016_J.Lipid.Res_57_131 |
| ESTHER : Iglesias_2016_J.Lipid.Res_57_131 |
| PubMedSearch : Iglesias_2016_J.Lipid.Res_57_131 |
| PubMedID: 26423520 |
| Title : Preparation of thioester substrates and development of continuous spectrophotometric assays for phospholipase A1 and monoacylglycerol lipase - Cox_1981_J.Lipid.Res_22_496 |
| Author(s) : Cox JW , Horrocks LA |
| Ref : J Lipid Res , 22 :496 , 1981 |
| Abstract : Cox_1981_J.Lipid.Res_22_496 |
| ESTHER : Cox_1981_J.Lipid.Res_22_496 |
| PubMedSearch : Cox_1981_J.Lipid.Res_22_496 |
| PubMedID: 7240974 |