1,2,3-trichloropropane
General
Type : Haloalkane
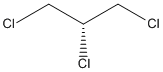
Chemical_Nomenclature : 1,2,3-trichloropropane
Canonical SMILES : C(C(CCl)Cl)Cl
InChI : InChI=1S\/C3H5Cl3\/c4-1-3(6)2-5\/h3H,1-2H2\/i1+1,2+1,3+1
InChIKey : CFXQEHVMCRXUSD-VMIGTVKRSA-N
Other name(s) : 1,2,3-Trichloropropane-13C3
MW : 150.4
Formula : C3H5Cl3
CAS_number :
PubChem :
UniChem :
Iuphar :
Target
Families : Haloalkane_dehalogenase-HLD2
References (7)
| Title : A Pseudomonas putida strain genetically engineered for 1,2,3-trichloropropane bioremediation - Samin_2014_Appl.Environ.Microbiol_80_5467 |
| Author(s) : Samin G , Pavlova M , Arif MI , Postema CP , Damborsky J , Janssen DB |
| Ref : Applied Environmental Microbiology , 80 :5467 , 2014 |
| Abstract : Samin_2014_Appl.Environ.Microbiol_80_5467 |
| ESTHER : Samin_2014_Appl.Environ.Microbiol_80_5467 |
| PubMedSearch : Samin_2014_Appl.Environ.Microbiol_80_5467 |
| PubMedID: 24973068 |
| Title : Crystallographic analysis of 1,2,3-trichloropropane biodegradation by the haloalkane dehalogenase DhaA31 - Lahoda_2014_Acta.Crystallogr.D.Biol.Crystallogr_70_209 |
| Author(s) : Lahoda M , Mesters JR , Stsiapanava A , Chaloupkova R , Kuty M , Damborsky J , Kuta Smatanova I |
| Ref : Acta Crystallographica D Biol Crystallogr , 70 :209 , 2014 |
| Abstract : Lahoda_2014_Acta.Crystallogr.D.Biol.Crystallogr_70_209 |
| ESTHER : Lahoda_2014_Acta.Crystallogr.D.Biol.Crystallogr_70_209 |
| PubMedSearch : Lahoda_2014_Acta.Crystallogr.D.Biol.Crystallogr_70_209 |
| PubMedID: 24531456 |
| Gene_locus related to this paper: rhoso-halo1 |
| Title : Substrate specificity of haloalkane dehalogenases - Koudelakova_2011_Biochem.J_435_345 |
| Author(s) : Koudelakova T , Chovancova E , Brezovsky J , Monincova M , Fortova A , Jarkovsky J , Damborsky J |
| Ref : Biochemical Journal , 435 :345 , 2011 |
| Abstract : Koudelakova_2011_Biochem.J_435_345 |
| ESTHER : Koudelakova_2011_Biochem.J_435_345 |
| PubMedSearch : Koudelakova_2011_Biochem.J_435_345 |
| PubMedID: 21294712 |
| Gene_locus related to this paper: agrtu-DHAA , brael-e2rv62 , braja-dhaa , myctu-linb , myctu-Rv1833c , rhoba-DHLA , rhoso-halo1 , sphpi-linb , xanau-halo1 |
| Title : Crystallization and crystallographic analysis of the Rhodococcus rhodochrous NCIMB 13064 DhaA mutant DhaA31 and its complex with 1,2,3-trichloropropane - Lahoda_2011_Acta.Crystallogr.Sect.F.Struct.Biol.Cryst.Commun_67_397 |
| Author(s) : Lahoda M , Chaloupkova R , Stsiapanava A , Damborsky J , Kuta Smatanova I |
| Ref : Acta Crystallographica Sect F Struct Biol Cryst Commun , 67 :397 , 2011 |
| Abstract : Lahoda_2011_Acta.Crystallogr.Sect.F.Struct.Biol.Cryst.Commun_67_397 |
| ESTHER : Lahoda_2011_Acta.Crystallogr.Sect.F.Struct.Biol.Cryst.Commun_67_397 |
| PubMedSearch : Lahoda_2011_Acta.Crystallogr.Sect.F.Struct.Biol.Cryst.Commun_67_397 |
| PubMedID: 21393851 |
| Gene_locus related to this paper: rhoso-halo1 |
| Title : Weak activity of haloalkane dehalogenase LinB with 1,2,3-trichloropropane revealed by X-Ray crystallography and microcalorimetry - Monincova_2007_Appl.Environ.Microbiol_73_2005 |
| Author(s) : Monincova M , Prokop Z , Vevodova J , Nagata Y , Damborsky J |
| Ref : Applied Environmental Microbiology , 73 :2005 , 2007 |
| Abstract : Monincova_2007_Appl.Environ.Microbiol_73_2005 |
| ESTHER : Monincova_2007_Appl.Environ.Microbiol_73_2005 |
| PubMedSearch : Monincova_2007_Appl.Environ.Microbiol_73_2005 |
| PubMedID: 17259360 |
| Gene_locus related to this paper: sphpi-linb |
| Title : Mechanism of enhanced conversion of 1,2,3-trichloropropane by mutant haloalkane dehalogenase revealed by molecular modeling - Banas_2006_J.Comput.Aided.Mol.Des_20_375 |
| Author(s) : Banas P , Otyepka M , Jerabek P , Petrek M , Damborsky J |
| Ref : J Comput Aided Mol Des , 20 :375 , 2006 |
| Abstract : Banas_2006_J.Comput.Aided.Mol.Des_20_375 |
| ESTHER : Banas_2006_J.Comput.Aided.Mol.Des_20_375 |
| PubMedSearch : Banas_2006_J.Comput.Aided.Mol.Des_20_375 |
| PubMedID: 17016745 |
| Title : Biodegradation of 1,2,3-trichloropropane through directed evolution and heterologous expression of a haloalkane dehalogenase gene - Bosma_2002_Appl.Environ.Microbiol_68_3582 |
| Author(s) : Bosma T , Damborsky J , Stucki G , Janssen DB |
| Ref : Applied Environmental Microbiology , 68 :3582 , 2002 |
| Abstract : Bosma_2002_Appl.Environ.Microbiol_68_3582 |
| ESTHER : Bosma_2002_Appl.Environ.Microbiol_68_3582 |
| PubMedSearch : Bosma_2002_Appl.Environ.Microbiol_68_3582 |
| PubMedID: 12089046 |
| Gene_locus related to this paper: rhoso-halo1 |